
前回説明した通り、炭疽病菌は葉の表に分生子層を作りますが、多くの炭疽病菌の分生子層には暗色で厚壁の剛毛が生えています(図1,2)。なぜでしょうか? その形は宿主の表皮を効率よく破って下に形成された分生子層を裸出させる役割を思い起こさせます。しかし、果樹などの多犯性炭疽病菌C. fioriniaeは硬い表皮の宿主にも分生子層を作りますが、それには剛毛がありません(図3,4)。また、何と言っても寒天培地上でできる分生子層にも剛毛はよく生えます。宿主上でなくても剛毛は生えるようにプログラムされているとも考えられますが、そんな無駄なことをするでしょうか。実は、その機能を考えるヒントとして培地上の剛毛形成条件が―その2―に出てきます。ということで表皮裂開説は当たっていないようです。
他の説は考えられないでしょうか? カビを食べる小動物は案外たくさんいます。クダアザミウマ亜目には胞子に穴をあけて中身を吸い取る食菌性の種がいますし(農林水産省,2017)、また、ダニ類には葉のダニ室(Domatium)に住み付いてうどんこ病菌を食べている種がいて、それを利用したブドウうどんこ病の生物防除が提案されました(Norton et al., 2000; Melidossian et al., 2005; English-Loeb, 2007)。炭疽病菌を専門に食べる小動物はまだ報告されていないと思われますが、分生子形成を促すために炭疽病の罹病葉を高湿度の容器に入れておくと、コナダニが侵入して分生子を食い荒らしてしまうことがあります。栄養の詰まった分生子やその形成細胞は食菌性の虫には魅力的な栄養源であり、炭疽病菌にとって繁殖器官を捕食されることは死活問題でしょう。食菌性のダニやトビムシなどは植物遺体を分解する菌類や細菌を主に食べるため落葉層から下に生息しています(横山,2015)。炭疽病菌が植物体上よりもむしろ地上に落ちた罹病茎葉・枝等の上で越冬する際、これらの虫に食害されるのは脅威です。ということで剛毛の役割は、食菌性小動物の攻撃に対する防御ではないでしょうか。既報の方法で準備した剛毛の有無だけが異なる分生子層に(Frost, 1964)、コナダニを放ち両分生子層の食害状況を調べればこの説が実験的に証明できるかもしれません。
ところで、剛毛を持たないかあるいは貧弱な剛毛を持つ種では捕食対策はどうしているのでしょうか? 剛毛のないC. acutatum群で柑橘類を加害する1種では生葉上の付着器で越冬し、翌春それが発芽して菌糸上に分生子を形成すると報告されています(佐藤,1997)。付着器は厚い頑丈な壁を持っており葉などに固着しているため、簡単には虫に食べられないと考えられます。
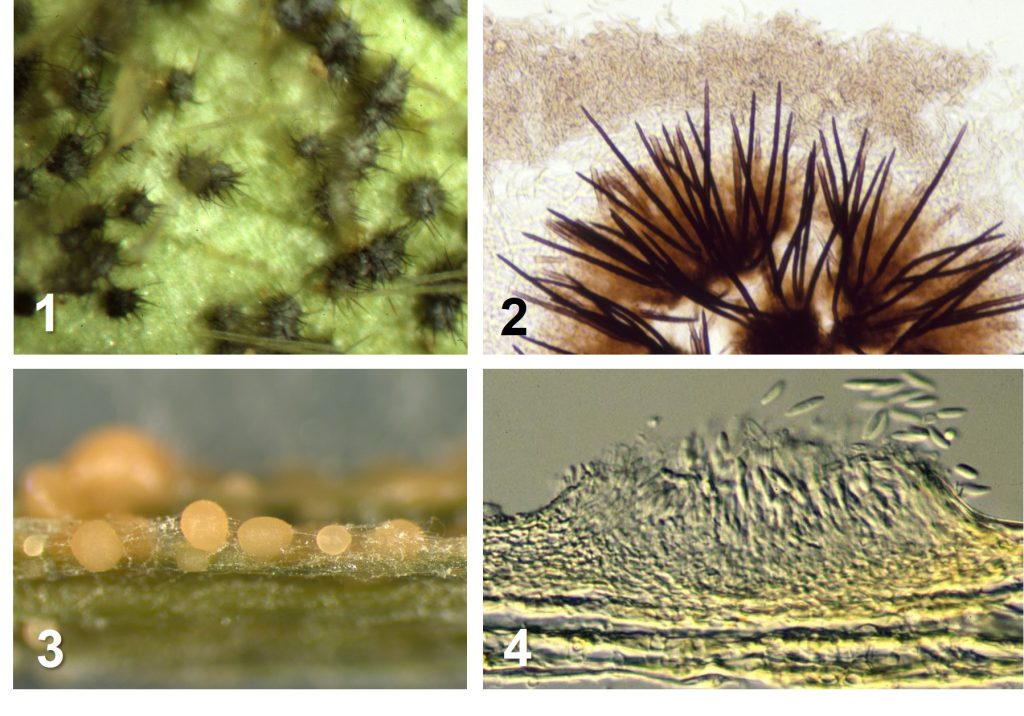
図 1と2はC. truncatum, 1. ダイズ莢上の分生子層の剛毛を示す.このうち,2はキク茎上の剛毛である。3と4はC. fioriniaeであり, 3は剛毛のない分生子層と分生子塊,4は分生子層の縦断面を示す。
引用文献
English-Loeb, G., Norton, A.P., Gadoury, D., Seem, R., Wilcox, W. 2007. Biological control of grape powdery mildew using mycophagous mites. Plant Dis. 91:421–429.
Frost, R.R. 1964. Seta formation in Colletotrichum spp. Nature 201: 730-731.
Melidossian H.S., Seem, R.C., English-Loeb, G., Wilcox, W.F. Gadoury, D.M. 2005. Suppression of grapevine powdery mildew by a mycophagous mite. Plant Dis. 89:1331–1338.
農林水産省 2017. 病害虫の同定に係る技術情報 -アザミウマ編(1)- 植物防疫所病害虫情報 第113 号 4pp.
Norton A.P., English-Loeb, G., Gadoury, D., Seem, R., 2000. Mycophagous mites and foliar pathogens: leaf domatia mediate tritrophic interactions in grapes. Ecology 81: 490-499.
佐藤豊三(1997):多犯性炭疽病菌 Colletotorichum acutatum の諸特性と同定法.四国植防 32:1-19.
横山和成 2015. 図解でよくわかる 土壌微生物のきほん.誠文堂新光社,東京 p. 118–119.